S. J. S. Rama Devi, Kuldeep Singh, B. Umakanth, B. Vishalakshi, P. Renuka, K. Vijay Sudhakar, M. S. Prasad, B. C. Viraktamath, V. Ravindra Babu, M. S. Madhav. .Development and Identification of Novel Rice Blast Resistant Sources and Their Characterization Using Molecular Markers. Chinese Journal of Rice Science,2015, 22(6): 300-308. 10.1016/S1672-6308(14)60308-5
Development and Identification of Novel Rice Blast Resistant Sources and Their Characterization Using Molecular Markers
S. J. S. Rama Devi1, Kuldeep Singh2, B. Umakanth1, B. Vishalakshi1, P. Renuka1, K. Vijay Sudhakar1, M. S. Prasad3, B. C. Viraktamath1, V. Ravindra Babu1, M. S. Madhav1
1Biotechnology, Crop Improvement, Indian Institute of Rice Research, Indian Council of Agriculture Research, Hyderabad 30, India
2School of Agricultural Biotechnology, Punjab Agricultural University, Ludhiana 141004, India
3Plant Pathology, Indian Institute of Rice Research, Indian Council of Agriculture Research, Hyderabad 30, India
To develop and characterize introgression lines for leaf and neck blast resistance, 326 introgression lines were developed using various accessions of six different AA genome wild species in the genetic background of elite Indian varieties like PR114 and Pusa 44 and were screened for blast resistance. Stringent phenotyping coupled with genotyping using gene based markers led to the identification of four resistant introgression lines, which showed promising resistance and do not possess any of the tested genes. Furthermore, multi-location screening confirmed the field resistance of the four introgression lines to both leaf and neck blast. Molecular characterization of these introgression lines using genome-wide simple sequence repeat markers revealed the presence of small percentage of wild Oryza genome introgrssion. So these lines can be used for mapping and identification of novel leaf and neck blast resistance genes. Thus, these four introgression lines can be considered as new genetic resources for blast resistance.
Keyword:
rice; blast resistance; introgression line; gene profiling; wild species
Rice is the staple food for more than half of the world’ s population, and global rice demand is estimated to rise from 6.76 × 108 t in 2010 to 8.52 × 108 t in 2035 (Khush, 2013). To produce 1.76 × 108 t additional rice, it is need to increase the yield and also minimize the yield loss caused by various diseases and insect pests. Rice blast disease, caused by Magnaporthe oryzae, is a major constraint for sustainable rice production. To date, around 100 blast resistance genes have been identified, and many of them have been cloned and characterized, such as Pb1, Pia, Pib, Pid2, Pid3, Pik, Pik-h/Pi54, Pik-m, Pik-p, Pish, Pit, Pita, Piz-t, Pi1, Pi2/Piz-5, Pi5, Pi9, Pi21, Pi25, Pi36, Pi37, Pi35, Pi64, Pi56, Pi63 and PiCO39 (Devanna et al, 2014). Though many resistant varieties to M. oryzae have been developed, the resistance is not long lasting, because the high pathogen plasticity in the fields makes single resistance gene break down after three to five years of the cultivar release (Bonman et al, 1986; Lang et al, 2009). Hence, development of broad spectrum and durable blast resistant varieties is essential for combating this disease, which requires continuous efforts of breeders and pathologists. Wild species of Oryzacan be exploited to widen the gene pool of rice for biotic and abiotic stress (Ram et al, 2013). Various studies have demonstrated that wild species as a reservoir of hidden gene(s) can be used for crop improvement like grassy stunt virus resistance gene fromO. nivara (Brar and Khush, 1997), QTLs for bacterial leaf blight and brown planthopper resistance (Zhang and Xie, 2014), and yield enhancing QTLs from various wild species (Linh et al, 2008; Rangel et al, 2008; Chen et al, 2009). Though wild species have been used extensively for other important agronomic traits of rice, they have been rarely used in blast resistance breeding programmes, since only two resistance genes, Pi9and Pi40, were reported from wild species (Amante et al, 1992; Jeung et al, 2007). Thus, it is worth to develop introgression lines (ILs) from various wild species and characterize for blast resistance. Further, stable ILs will facilitate the use of wild species derived genes/alleles in breeding programmes (Lei et al, 2013; Zhou et al, 2014). The objective of this study was to identify novel genetic resources for blast disease resistance. Here we reported development and systematic screening of ILs across India for field resistance to rice blast. Since, field resistance allows effective control of a pathogen under natural field conditions and it is considered to be durable when exposed to new races of blast (Fukuoka and Okuno, 2001). We also made an attempt to identify the reported blast resistance (R) genes among the resistant ILs using gene based markers which are reported earlier (Hayashi et al, 2006; Lmam et al, 2014). In addition to the identification of blast R genes, we tried to identify the introgressed regions among the identified ILs using genome-wide simple sequence repeat (SSR) markers. SSR markers are most popular genetic markers with high polymorphism rate, high abundance and broad distribution throughout the genome, which are inherited in a Mendelian fashion as codominant markers (Miah et al, 2013). Also, the availability of many SSR markers in rice helps the breeders for crop improvement through molecular marker assisted breeding (McCouch et al, 2002). The strategic phenotyping for blast disease (field) resistance coupled with genotyping using markers has led to the identification of resistant ILs which are valuable genetic resources not only for blast resistance but also for novel gene identification leading to cultivar development (Ashikari and Matsuoka, 2006; Rahman et al, 2011).
MATERIALS AND METHODS
Development of introgression lines
The stable ILs was developed at Punjab Agricultural University, Ludhiana, India, with PR114 and Pusa 44 as recurrent parents and six wild species of Oryza (AA genome) as donors. These lines were developed through repeated backcrossing coupled with phenotypic selection for yield parameters. The donor species included 30 accessions of O. glaberrima, O. nivara, O. rufipogon, O. longistaminata, O. glumaepatulaandO. barthii(Supplemental Table 1). PR114 and Pusa 44 are the elite rice varieties with high yield and good cooking quality (Gaikwad et al, 2014).
Rice materials
From the 326 rice ILs (BC2-3F9) of PR114 and Pusa 44, single plant was selected based on the maximum features of its recurrent parent, and the seeds of single plant were used for further studies. Six monogenic lines carrying single blast R gene and one variety harbouring the rice blast R genes were used as resistant checks for identification of known gene allele among the four ILs (Table 1). The varieties viz, LTH, BPT5204, HR12 and Co39 were used as susceptible checks for gene profiling.
Table 1
Table 1
Table 1 List of blast resistant genes tested among introgression lines using gene based markers.
Gene
Variety
Chr
Marker
Size (kb)
Forward primer (5′ -3′ )
Reverse sequence (5′ -3′ )
Reference
Piz-t
IRBLb-B[LT]
6
Z56592
292
ggacccgcgttttccacgtgtaa
aggaatctattgctaagcatgac
Hayashi et al, 2006
Pita/
IRBLta-Zh[LT]
12
Pita3
861
agtcgtgcgatgcgaggacagaaac
gcattctccaacccttttgcatgcat
Imam et al, 2014
Pita2
IRBLta2-p1[LT]
12
YL155/YL87
1042
agcaggttataagctaggcc
ctaccaacaagttcatcaaa
Imam et al, 2014
Pi9
IRBL9-W[LT]
6
195R-1
2000
Atggtcctttatctttattg
ttgctccatctcctctgtt
Qu et al, 2006
6
NMSMPi9-1
168
cgagaaggacatctggtacg
gagatgcttggatttagaagac
Qu et al, 2006
Pi54
Tetep
11
TRS26
266
ggagagccaatctgataagca
caacaagagaggcaaattctca
Sharma et al, 2005
11
Pikh MAS
216
caatctccaaagttttcagg
gcttcaatcactgctagacc
Ramkumar et al, 2011
Pi40
IR65482-4-1-136-2-2
6
MSM6
256
tgctgagatagccgagaaatc
gcacccttttcgctagagg
Rama Devi et al, 2013
6
9871.T7E2b
641
caacaaacgggtcgacaaagg
cccccaggtcgtgataccttc
Jeung et al, 2007
Pi1
C101LAC
11
RM224
157
atcgatcgatcttcacgagg
tgctataaaaggcattcggg
Hittalmani et al, 2000
11
RM1233
170
aataggcctggagagaatttcc
ccttataagccgtctcgatcc
Fuentes et al, 2008
Pi2
C101A51
6
MSM1
175
gctagtgaagcaattcctatgg
caagaaaatggccagaacg
Arunakanthi et al, 2008
6
AP56595
288
Ctccttcagctgctcctc
tgatgacttccaaacggtag
Fjellstrom et al, 2004
Chr, Chromosome.
Table 1 List of blast resistant genes tested among introgression lines using gene based markers.
Fungal inoculation and evaluation of field resistance to blast
The seeds of ILs were sown on uniform blast nursery beds along with local susceptible checks HR12 (tall, low tillering variety with susceptibility to leaf blast), Co39 (universal blast susceptible variety), and BPT5204 (popularly known as Samba Mahsuri, mega variety with high yield and good quality, but susceptible to rice blast). Among the three checks, HR12 was used as spreader row. After 15 d of germination, the plants were inoculated with most virulent isolates of India i.e. NLR-1 maintained as DRR (Directorate of Rice Research) isolate (Padmavathi et al, 2005; Prasad et al, 2009; Ratna et al, 2011). The inoculum contained 50 000 conidia/mL with 0.5% glycerol and the nursery beds were maintained with 90% relative humidity. The ILs was extensively screened for blast resistance as follows: (i) Screened for resistance at the Indian Institute of Rice Research in three seasons (January to April in 2010 and 2011, and June to September in 2011), with three replications in each season; (ii) The selected ILs was screened at seven blast endemic areas to know their leaf blast resistance in the two seasons (June to September 2011 and 2012); (iii) The short listed ILs were further screened at hilly areas (which are hotspots for natural neck blast infestation in India) to check their resistance to neck blast (June to September in 2011 and 2012) (Supplemental Table 2); and (iv) To check the field resistance, the selected ILs were screened across different parts of the country through All India Co-ordinated Rice Improvement Programme (AICRIP) in 2012 and 2013 (http://www.drr icar.org/aicrip.htm). In AICRIP screening, the ILs were screened at 27 different centres for leaf blast and 8 centres for neck blast. The blast screenings of (ii), (iii) and (iv) were conducted in two replications per season and the mean scores were obtained. All the scores were obtained according to the standard evaluation system (Supplemental Table 3) (Panguluri et al, 2013). The selected four ILs were evaluated for the essential agronomic traits in the fields. The plants were grown at Indian Institute of Rice Research during Kharif in 2012 and 2013. Individual plants from the ILs were evaluated and the data for plant height, number of tillers per plant, number of panicles per plant, yield per plant and biomass were documented from 10 plants for each entry and the mean values were determined.
DNA extraction and identification of blast resistance genes
Genomic DNA was isolated using modified cetyltrimethyl ammonium bromide (CTAB) method (Saghai et al, 1984) and the quality of isolated DNA was checked using 0.8% agarose gel electrophoretically and quantified using the nanodrop (Thermo Fisher, USA). To identify the presence of blast resistance gene alleles among the ILs, gene profiling was carried out according to Lmam et al (2014). For this, seven effective blast resistance genes (Pi54, Piz-t, Pi1, Pi9, Pi2, Pita/Pita2and Pi40) were analyzed among blast resistant ILs using gene based markers (for Pi54, Piz-t, Pi9, Pita/Pita2, Pi40andPi2) as well as two flanking markers (for Pi1) (Table 1). PCR was performed using 1 U of Taq DNA polymerase and 10 × PCR buffer (Genei, India) in 15-µ L reaction volume with a thermal profile as described in Lmam et al (2014). The PCR products were resolved on agarose gel (4% for the products less than 500 bp and 2% for the products more than or equal to 1 kb) in 0.5 × TBE buffer, stained with ethidium bromide (0.5 pg/mL) and documented under UV light. Scoring was done based on presence (1) or absence (0) of the gene in comparison with the resistant and susceptible checks.
Marker analysis
The four ILs (IL-1 to IL-4) were forwarded further for precise identification of the donor genome introgression. A set of 499 SSR markers (McCouch et al, 2002) which were spread uniformly across the twelve linkage groups were used. Scoring of the ILs was done by comparing the IL alleles with the recurrent parent allele and the donor allele.
In silico analysis of target region
The introgressed regions in the four ILs were analyzed based on the existing literature for the presence of meta QTLs (Ballini et al, 2008). Further, in silico analysis of the introgressed region was done by using RAP-DB (http://rapdb.dna.affrc.go.jp/). For this, the list of annotated genes between the introgressed regions were downloaded from the database and categorized them based on their molecular functions. Later the genes associated with disease resistance were listed.
RESULTS
Development of introgression lines
The ILs were developed through backcrossing followed by recurrent selection. In brief, two or three backcrosses were carried out between donor and recurrent parents (PR114 and Pusa 44). After each backcross, the plants were selected based on phenotypic characters as well as yield related traits which were close to the recurrent parent. This is followed by continuous selfing so as to develop BC2-3F9 generation, at this stage, almost all the ILs were closely resemble to their respective recurrent parents. The process of developing the ILs is summarized in Supplemental Fig. 1.
Fig. 1. Evaluation of blast disease at Directorate of Rice Research, India, during 2010 and 2011. (A, B, C and D are IL-1, IL-2, IL-3 and IL-4 showing immune response with susceptible check HR12 on either side, respectively.)
Blast screening
The 326 ILs were screened for leaf blast resistance at DRR in three seasons (2010 to 2011), 50 ILs showed resistant reaction with mean score at 0 to 3 (Fig. 1 and Supplemental Table 4) and 276 were susceptible with mean score at 4 to 9), while both the recipient parents showed extreme susceptibility. Among the 50 resistant ILs, surprisingly majority of the lines (41) were under the PR114 background and only 9 lines were under the Pusa 44 background. Among all the wild species screened for blast, O. glaberrimacontributed major share (34%) of resistance, followed by O. longistaminata(32%), O. nivara, O. rufipogon, O. glumaepatula (each species for 10%) while the least fromO. barthii(4%). Among the 50 resistant ILs, top four ILs (designated as IL-1 to IL-4, Table 2) were selected (based on the phenotypic data ranging from 1.0 to 1.5) for screening at endemic regions of leaf and neck blast to check their field resistance across different parts of India (Supplemental Table 2). All the ILs showed high level of resistance, and their mean resistance score were in the range of 1.3 to 2.6. IL-1 showed the higher resistance (mean score of 1.3) than the others (Table 2). Furthermore, the screening at hilly areas for neck blast resistance indicated that all the lines were resistant to neck blast with the mean scores ranging from 1.6 to 2.3 (Table 2). These ILs screened for multiple disease resistance under AICRIP during 2012 and 2013 showed leaf and neck blast resistance with susceptibility index ranging from 3.3 to 3.8 and 2.0 to 3.0, respectively. The comparison of blast scores were also made with C101LAC (monogenic line for Pi1), C101A51 (monogenic line for Pi2) and HR12 (local susceptible check). The results showed that the ILs showed resistant reaction better than those of lines containing Pi1 and Pi2 genes, the resistant checks (Table 2). The agronomic trait evaluation of the top four ILs has revealed that the ILs performance was at par with that of the recurrent parents (Supplemental Table 5 and Supplemental Fig. 2).
Table 2
Table 2
Table 2 Evaluation of introgression lines for leaf and neck blast resistance.
Code
Detail of cross a
Leaf blast b
Neck blast b
IIRR
India c
AICRIP d
Mean
SD
Variance
India c
AICRIP d
Mean
SD
Variance
IL-1
PR114/O. glumaepatula(104387)//2× PR114
1
1.3
3.5
1.9
1.4
1.86
1.6
2.9
2.3
0.9
0.9
IL-2
PR114/O. glaberrima (102526)//3× PR114
1
1.4
3.3
1.9
1.2
1.51
1.8
2.9
2.4
0.8
0.6
IL-3
PR114/O. nivara(105410)//2× PR114
1.2
2.6
3.5
2.4
1.2
1.34
2.3
2
2.2
0.2
0.1
IL-4
Pusa 44/O.barthii (101248)//3× Pusa 44
1.3
2.4
3.8
2.5
1.3
1.57
2.3
3
2.7
0.5
0.3
C101LAC e
Monogenic line of Pi1
1.6
2.1
3.6
2.4
1
1.08
2.9
3.5
3.2
0.4
0.2
C101A51 e
Monogenic line of Pi2
1.6
2
4.3
2.6
1.5
2.12
3.1
3.9
3.5
0.6
0.3
HR12 f
9
8
6.3
7.8
1.4
1.86
6.4
5.8
6.1
0.4
0.2
a The number in the parentheses is the accession number and the number before ‘ × ’ means the times of backcross; b Mean phenotypic score (scale from 0 to 9 according to the standard evaluation system) recorded; c Mean phenotypic score obtained from hotspots of India (seven centers for leaf blast and five centers for neck blast); d Mean phenotypic score obtained from All India Co-ordinated Rice Improvement Programme (AICRIP) in 2012 and 2013 (27 centers for leaf blast and 8 centers for neck blast); e Resistant check; f Susceptible check.
IIRR, Indian Institute of Rice Research; SD, Standard deviation.
Table 2 Evaluation of introgression lines for leaf and neck blast resistance.
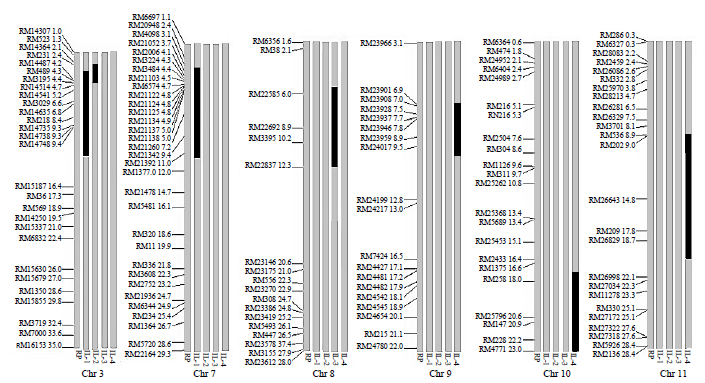
Fig. 2. Determination of donor genome introgression in four introgression lines (ILs) which is made by comparing ILs with recurrent parent.(Chr, Chromosome. The black region represents the donor introgression from the wild species.)
Identification of blast resistance genes using gene based markers
To know the allelic status of gene in the ILs, genotyping was done using gene based markers associated with blast R genes. The ILs which showed marker alleles same to positive check were considered as positive for the gene (score as 1), similarly, the ILs which showed marker allele correlating to the susceptible checks were considered as negative for the gene (score as 0). A total of seven important blast resistance genes of India were genotyped in the present study, and none of these four ILs showed the presence of tested blast R gene alleles (Supplemental Table 6).
Determination of introgression region in introgression lines
To identify the introgressed genomic regions from the wild Oryza species, four ILs were screened using 499 SSR markers (selected based on at least one marker per Mb) (Fig. 2). The results revealed the presence of 0.6% to 4.6% donor genome introgression (Supplemental Fig. 3). Most introgressions were observed at one region of particular chromosome except in IL-1 and IL-4. These two ILs showed the introgression at multiple chromosomes. The physical distances of introgressed regions among ILs varied from 2.3 to 9.0 Mb across different chromosomes (Table 3 and Supplemental Fig. 4). IL-2 had the minimum introgression (0.6%) of donor genome at chromosome 3 from 2.4 to 4.3 Mb region whereas IL-4 had the maximum introgression (4.6%) (Table 4). This information will facilitate in precise mapping and identification of gene(s) in these ILs.
In silico analysis of target region
The meta QTL analysis revealed the presence of meta QTLs in all four ILs where the donor genome introgression regions were found. In IL-3 and IL-4, we determined the presence of blast R genes. Through in silico, we observed the presence of nucleotide binding site-leucine rich repeat, nucleotide-binding adaptor shared by ARC domain containing protein, and serine/ threonine kinase genes in the target regions in all the ILs, which may be the candidate genes involved in resistance (Table 3).
Table 3
Table 3
Table 3 Details of in silico analysis of introgression regions of four resistant introgression lines.
Table 3 Details of in silico analysis of introgression regions of four resistant introgression lines.
Table 4
Table 4
Table 4 Molecular analysis for determination of introgression among four introgression lines (ILs).
IL
No. of polymorphic markers on each chromosome
Total number of polymorphic markers
Percentage of recurrent genome
Percentage of donor genome
Chr 1
Chr 2
Chr 3
Chr 4
Chr 5
Chr 6
Chr 7
Chr 8
Chr 9
Chr 10
Chr 11
Chr 12
(%)
(%)
-26
-39
-60
-55
-27
-68
-72
-44
-26
-26
-26
-30
IL-1
0
0
10
0
0
0
13
0
0
0
0
0
23
95.4
4.61
IL-2
0
0
3
0
0
0
0
0
0
0
0
0
3
99.4
0.6
IL-3
0
0
0
0
0
0
0
4
0
0
0
0
4
99.2
0.8
IL-4
0
0
0
0
0
0
0
0
6
5
3
0
14
97.2
2.8
The values in the parentheses are the number of markers tested on each chromosome (Chr), respectively.
Table 4 Molecular analysis for determination of introgression among four introgression lines (ILs).
DISCUSSION
Wild species are rich resources for a variety of important genes which have agronomical importance (Cheema et al, 2008; Nataraj et al, 2011) including disease resistance (Jena et al, 2006; Ramkumar et al, 2011). However, till today only two genes (Pi9 and Pi40) have been identified from wild species (Amante et al, 1992; Jeung et al, 2007). ILs have proven to be potential genetic resources for detecting rice blast resistance (Tsunematsu et al, 2000; Rahman et al, 2011) and also for exploring the agronomic traits (Fujita et al, 2009; Gu et al, 2012). Since we used different AA genome for the development of the ILs, it is expected to identify novel genetic resources for rice blast disease. Hence it is worth screening ILs for blast resistance, upon stringent screening at DRR with most virulent race in artificial screening conditions, 50 ILs were resistant and quite numbers of ILs were susceptible (Supplemental Table 1), since the advancement of generations of these lines were done earlier based on the agro-morphological and yield-related traits which resembles the recurrent parent. Majority of the resistant ILs were derived from O. glaberrima and O. longistaminata. Recently, Sié et al (2012) and He et al (2014) have demonstrated the utility of O. longistaminataand O. glaberrima for many valuable traits like rhizomatousness, disease resistance and drought tolerance that can be used to improve cultivated rice through transcriptomic, proteomic and metabolomic studies, and they exploited the several accessions of these wild species in breeding programs. Out of 50, only 4 lines were further advanced to check their field resistance at multiple locations, which included hotspots for leaf and neck blast. These hotspots, known to have diverse pathogen populations, generally offer field resistance, which is much better than testing with the single isolates of the pathogen. Further, these lines were also tested with artificial and natural infestations with predominant blast isolates of various test sites of AICRIP programme. The tested four ILs showed consistently resistant reactions in all the tested locations and seasons. Though there are many reports on the leaf blast resistance genes, less information is known towards neck blast, as only one gene Pb1 has been reported so far (Hayashi et al, 2010). In this context, the four ILs can be used as potential donors for neck blast resistance breeding programmes in India. The top four ILs were derived from O. glumaepatula (IL-1), O. glaberrima (IL-2), O. nivara (IL-3) and O. barthii(IL-4), respectively. Very interestingly, till date no major gene or major QTL has been identified from any of the four wild species from which the ILs have derived. The rest 46 resistant ILs were unexplored but the material is worth enough to look into further.
Apart from stringent phenotypic screening, the gene based markers were used to identify the known blast R alleles (Hayashi et al, 2006; Lmam et al, 2014). For this study, seven blast R genes which are being used more often in blast resistance breeding programs in India were selected (Prasad et al, 2011). Moreover, it is difficult and time-consuming process to conduct allelism tests for all the known blast R genes. Present study revealed that the selected four ILs does not have alleles of the seven tested blast R genes. Similar approach was followed earlier for identification of alleles or better performing alleles from the diverse germplasm through PCR based approach (Mahender et al, 2012; Ingole Kishor et al, 2014; Khan et al, 2014; Lmam et al, 2014).
Availability of large number of reliable SSR markers in rice and the availability of donor accessions, we can precisely locate the donor genome introgression of four ILs. Interestingly, the donor genome introgression was small in most of the ILs and these genomic regions were not reported earlier for presence of major blast resistance genes (at least in two ILs), indicating the presence of novel genes. Increasing the marker density along the chromosomes will enhance the precise identification of introgression regions. In the present study, 499 SSR markers (one marker per Mb) spread uniformly across the genome (except on chromosomes 8 and 9) were selected for this assay. Further increase in density of markers would certainly aid in the narrowing down the introgression regions. This observation was further strengthened by in silico analysis, which showed the presence of various disease related genes in the introgressed regions among the ILs (Table 3). In addition, we observed the presence of meta QTLs in the introgressed regions of all ILs (Ballini et al, 2008), indicating the importance of these genomic regions in the point of field resistance. This information will be useful in mapping of genes associated with resistance. All the ILs used in this study were stable lines, since they were in BC2-3F9 generation which can directly enter into the cultivar development programme. These lines can serve as good resources for the identification of novel genes for leaf and neck blast resistance. Two of these lines (IL-1 and IL-2) were registered (INGR15001 and INGR15002) by the National Bureau of Plant Genetic Resources (NBPGR), India, which is a responsible organization for repository and proper distribution of the ILs as open materials for further studies. Further, we identified that 46 ILs which are also resistant to blast and multiple disease, can be further evaluated for other biotic and abiotic stress. To conclude, we used the wild species in resistance breeding programmes by developing ILs, and we followed systemic approach to identify the ILs which showed the stable blast resistance. The scope of this study lies in the identification of four valuable and novel genetic resources which can be exploited further for blast resistance breeding programmes.
ACKNOWLEDGEMENTS
S. J. S. Rama Devi thanks the Council for Scientific and Industrial Research (CSIR), New Delhi, India and Department of Genetics, Osmania University, Hyderbad, Inida for providing Senior Research Fellowship.
SUPPLEMENTAL DATA
The following materials are available in the online version of this article at http://www.sciencedirect.com/science/ journal/16726308; http://www.ricescience.org.
Supplemental Table 1. List of introgression lines used in this study.
Supplemental Table 2. List of leaf and neck blast hot spot areas in India.
Supplemental Table 3. Disease severity score and corresponding symptoms on plants.
Supplemental Table 4. Resistant introgression lines identified at Indian Institute of Rice Research.
Supplemental Table 5. Agronomic evaluation of the introgression lines.
Supplemental Table 6. Identification of rice blast resistance genes using gene specific markers.
Supplemental Fig. 1. Development and blast disease resistance evaluation of introgression lines.
Supplemental Fig. 2. Agronomic trait evaluation of the four introgression lines.
Supplemental Fig. 3. Representative gel pictures for identification of donor genome in introgression lines IL-1, IL-2 and IL-3.
Supplemental Fig. 4. Precise identification of donor genome introgression in selected introgression lines using genome wide simple sequence repeat markers.
The authors have declared that no competing interests exist.
Amante BA, Sitch LA, NelsonR, Dalmacio RD, Oliva NP, AswidinnoorH, LeungH. 1992. Transfer of bacterial blight and blast resistance from the tetraploid wild rice Oryza minuta to cultivated rice, Oryza sativa. Theor Appl Genet, 84(3/4): 345-354. [Cited within:2]
[2]
ArunakanthiB, Prasad MS, MadhanmohanK, Balachand ran SM, Madhav MS, Reddy CS, Viraktamath BC. 2008. Introgression of major blast resistance genes Pi-1, Pi-2 and Pi-kh in indica rice cultivars Samba Mahsuri and Swarna. J Mycol Plant Pathol, 38(3): 625-630. [Cited within:1]
[3]
AshikariM, MatsuokaM. 2006. Identification, isolation and pyramiding of quantitative trait loci for rice breeding. Trends Plant Sci, 11(7): 344-350. [Cited within:1]
[4]
BalliniE, Morel JB, DrocG, PriceA, CourtoisB, Notteghem JL, TharreauD. 2008. A genome-wide meta-analysis of rice blast resistance genes and quantitative trait loci provides new insights into partial and complete resistance. Mol Plant Microbe Interact, 21(7): 859-868. [Cited within:2]
[5]
BerruyerR, AdreitH, MilazzoJ, GaillardS, BergerA, DiohW, Lebrun MH, TharreauD. 2003. Identification and fine mapping of Pi33, the rice resistance gene corresponding to the Magnaporthe grisea avirulence gene ACE1. Theor Appl Genet, 107(6): 1139-1147. [Cited within:1]
[6]
Bonman JM, Dedios T IV, Khin MM. 1986. Physiological specialization of Pyricularia oryzae in the Philippines. Plant Dis, 70(8): 767-769. [Cited within:1]
[7]
Brar DS, Khush GS. 1997. Alien introgression in rice. In: Oryza: From Molecule to Plant. Netherland s: Springer: 10-47. [Cited within:1]
[8]
Cheema KK, Grewal NK, VikalY, SharmaR, Lore JS, DasA, BhatiaD, MahajanR, GuptaV, Bharaj TS, SinghK. 2008. A novel bacterial blight resistance gene from Oryza nivara mapped to 38 kb region on chromosome 4L and transferred to Oryza sativa L. Genet Res, 90(5): 397-407. [Cited within:1]
[9]
Chen ZW, Hu FY, XuP, LiJ, Deng XN, Zhou JW, LiF, Chen SN, Tao DY. 2009. QTL analysis for hybrid sterility and plant height in interspecific populations derived from a wild rice relative, Oryza longistaminata. Breeding Sci, 59: 441-445. [Cited within:1]
[10]
Devanna NB, VijayanJ, Sharma TR. 2014. The blast resistance gene Pi54of cloned from Oryza officinalis interacts with Avr-Pi54 through its novel non-LRR domains. PLoS One, 9(8): e104840. [Cited within:1]
[11]
FjellstromR, McClung AM, Conaway-Bormans CA, Anna MM, Marchetti MA, Shank AR, Park WD. 2004. Development of DNA markers suitable for marker assisted selection of three Pi genes conferring resistance to multiple Pyricularia grisea pathotypes. Crop Sci, 44(5): 1790-1798. [Cited within:1]
[12]
FjellstromR, McClung AM, Shank AR. 2006. SSR markers closely linked to the Pi-z locus are useful for selection of blast resistance in a broad array of rice germplasm. Mol Breeding, 17(2): 149-157. [Cited within:1]
[13]
Fuentes JL, Correa-Victoria FJ, EscobarF, PradoG, AricapaG, Duque MC, TohmeJ. 2008. Identification of microsatellite markers linked to the blast resistance gene Pi1(t) in rice. Euphytica, 160: 295-304. [Cited within:1]
[14]
FujitaD, Santos RE, Ebron LA, Telebanco-Yanoria MJ, KatoH, KobayashiS, UgaY, ArakiE, TakaiT, TsunematsuH, ImbeT, Khush GS, Brar DS, FukutaY, KobayashiN. 2009. Development of introgression lines of an indica-type rice variety, IR64, for unique agronomic traits and detection of the responsible chromosomal regions. Field Crops Res, 114(2): 244-254. [Cited within:1]
[15]
FukuokaS, OkunoK. 2001. QTL analysis and mapping of pi21, a recessive gene for field resistance to rice blast in Japanese upland rice. Theor Appl Genet, 103(2/3): 185-190. [Cited within:1]
[16]
FukuokaS, Yamamoto SI, MizobuchiR, YamanouchiU, OnoK, KitazawaN, YasudaN, FujitaY, Nguyen T TT, KoizumiS, SugimotoK, MatsumotoT, YanoM. 2014. Multiple functional polymorphisms in a single disease resistance gene in rice enhance durable resistance to blast. Sci Rep, 4: 4550. [Cited within:1]
[17]
Gaikwad KB, SinghN, BhatiaD, KaurR, Bains NS, Bharaj TS, SinghK. 2014. Yield-enhancing heterotic QTL transferred from wild species to cultivated rice Oryza sativa L. PLoS One, 9(6): e96939. [Cited within:1]
[18]
GowdaM, Barman SR, Chattoo BB. 2006. Molecular mapping of a novel blast resistance gene Pi38 in rice using SSLP and AFLP markers. Plant Breeding, 125(6): 596-599. [Cited within:1]
[19]
GuJ, YinX, Stomph TJ, WangH, Struik PC. 2012. Physiological basis of genetic variation in leaf photosynthesis among rice (Oryza sativa L. ) introgression lines under drought and well-watered conditions. J Exp Bot, 63(14): 5137-5153. [Cited within:1]
[20]
HayashiK, YoshidaH, AshikawaI. 2006. Development of PCR-based allele-specific and InDel marker sets for nine rice blast resistance genes. Theor Appl Genet, 113(2): 251-260. [Cited within:2]
[21]
HayashiN, InoueH, KatoT, FunaoT, ShirotaM, ShimizuT, KanamoriH, YamaneH, HayanosaitoY, MatsumotoT, YanoM, TakatsujiH. 2010. Durable panicle blast-resistance gene Pb1 encodes an atypical CC-NBS-LRR protein and was generated by acquiring a promoter through local genome duplication. Plant J, 64(3): 498-510. [Cited within:1]
[22]
He RF, SalvatoF, Park JJ, Kim MJ, NelsonW, Balbuena TS, WillerM, Crow JA, May GD, Soderlund CA, Thelen JJ, Gang DR. 2014. A systems-wide comparison of red rice (Oryza longistaminata) tissues identifies rhizome specific genes and proteins that are targets for cultivated rice improvement. BMC Plant Biol, 14: 46. [Cited within:1]
[23]
HittalmaniS, ParcoA, Mew TV, Zeigler RS, HuangN. 2000. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice. Theor Appl Genet, 100(7): 1121-1128. [Cited within:1]
[24]
ImamJ, AlamS, Mand al NP, VariarM, ShuklaP. 2014. Molecular screening for identification of blast resistance genes in North East and Eastern Indian rice germplasm (Oryza sativa L. ) with PCR based makers. Euphytica, 196(2): 199-211. [Cited within:1]
[25]
Ingole KishorD, Prashanthi SK, Krishnaraj PU. 2014. Mining for major blast resistance genes in rice land races of Karnataka. Ind J Genet Plant Breeding, 74(3): 378-383. [Cited within:1]
[26]
Jena KK, Jeung JU, Lee JH, Choi HC, Brar DS. 2006. High-resolution mapping of a new brown planthopper (BPH) resistance gene, Bph18(t), and marker-assisted selection for BPH resistance in rice (Oryza sativa L. ). Theor Appl Genet, 112(2): 288-297. [Cited within:1]
[27]
Jeung JU, Kim BR, Cho YC, Han SS, Moon HP, Lee YT, Jena KK. 2007. A novel gene, Pi40(t), linked to the DNA markers derived from NBS-LRR motifs confers broad spectrum of blast resistance in rice. Theor Appl Genet, 115(2): 1163-1177. [Cited within:2]
[28]
Khan M AI, Sen PP, BhuiyanR, KabirE, Chowdhury AK, FukutaY, AliA, Latif MA. 2014. Phenotypic screening and molecular analysis of blast resistance in fragrant rice for marker assisted selection. Compt Rend Biol, 337(5): 318-324. [Cited within:1]
[29]
Khush GS. 2013. Strategies for increasing the yield potential of cereals: Case of rice as an example. Plant Breeding, 132(5): 433-436. [Cited within:1]
[30]
Lang NT, Luy TT, Ha P TT, Buu BC. 2009. Monogenic lines resistance to blast disease in rice (Oryza sativa L. ) in Vietnam. Int J Genet Mol Biol, 1(7): 127-136. [Cited within:1]
[31]
Lei MP, Li GR, ZhouL, Li CH, LiuC, Yang ZJ. 2013. Identification of wheat-Secale africanum chromosome 2Rafr introgression lines with novel disease resistance and agronomic characteristics. Euphytica, 194(2): 197-205. [Cited within:1]
[32]
Linh LH, Hang NT, Jin FX, Kang KH, Lee YT, Kwon SJ, Ahn SN. 2008. Introgression of a quantitative trait locus for spikelets per panicle from Oryza minuta to the O. sativa cultivar Hwaseongbyeo. Plant Breeding, 127(3): 262-267. [Cited within:1]
[33]
LmamJ, AlamS, Mand al NP, VariarM, ShuklaP. 2014. Molecular screening for identification of blast resistance genes in North East and Eastern Indian rice germplasm (Oryza sativa L. ) with PCR based makers. Euphytica, 196(2): 199-211. [Cited within:3]
[34]
MahenderA, Swain DM, Subudhi HN, Rao G JN. 2012. Molecular analysis of native Manipur rice accessions for resistance against blast. Afr J Biotechnol, 11(6): 1321-1329. [Cited within:1]
[35]
McCouch SR, TeytelmanL, XuY, Lobos KB, ClareK, WaltonM, FuB, MaghirangR, LiZ, XingY, ZhangQ, KonoI, YanoM, FjellstromR, De ClerckG, SchneiderD, CartinhourS, WareD, SteinL. 2002. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L. ). DNA Res, 9(6): 199-207. [Cited within:2]
[36]
MiahG, Rafii MY, Ismail MR, Puteh AB, Rahim HA, Islam KN, Latif MA. 2013. A review of microsatellite markers and their applications in rice breeding programs to improve blast disease resistance. Int J Mol Sci, 14: 22499-22528. [Cited within:1]
[37]
Nataraj KumarP, SujathaK, Laha GS, Srinivasa RaoK, MishraB, Viraktamath BC, HariY, Reddy CS, Balachand ran SM, RamT, Sheshu MadhavM, Shobha RaniN, Neeraja CN, Ashok ReddyG, ShaikH, Sundaram RM. 2011. Identification and fine- mapping of Xa33, a novel gene for resistance to Xanthomonas oryzae pv oryzae. Phytopathology, 102(2): 222-228. [Cited within:1]
[38]
PadmavathiG, RamT, SatyanarayanaK, MishraB. 2005. Identification of blast (Magnaporthe grisea) resistance genes in rice. Curr Sci, 88(4): 628-630. [Cited within:1]
[39]
Panguluri SK, Kumar AA. 2013. Phenotyping for Plant Breeding. New York: Springer: 1-40. [Cited within:1]
[40]
Prasad MS, Kanthi BA, Balachand ran SM, SeshumadhavM, Mohan KM, Viraktamath BC. 2009. Molecular mapping of rice blast resistance gene Pi-1(t) in the elite indica variety Samba Mahsuri. World J Microbiol Biotechnol, 25(10): 1765-1769. [Cited within:1]
QuS, LiuG, ZhouB, BellizziM, ZengL, DaiL, HanB, Wang GL. 2006. The broad spectrum blast resistance gene Pi9 encodes a nucleotide-binding site leucine-rich repeat protein and is a member of a multigene family in rice. Genetics, 172: 1901-1914. [Cited within:1]
[43]
RahmanL, KhanamS, JaehwanR, HeejongK. 2011. Mapping of QTLs Involved in resistance to rice blast (Magnaporthe grisea) using Oryza minuta introgression lines. Czech J Genet Plant Breeding, 47(3): 85-94. [Cited within:2]
[44]
RamT, Bhadana VP, Laha GS, Padmakumari AP, Jyothibadri, Azam MM, Amtulwaris, SarlaN, PadmavathiG, DineshC, DivyaB, Viraktamath BC. 2013. Wild Species in Rice Improvement. Hyderabad, India: Directorate ofRice Research. [Cited within:1]
[45]
Rama Devi S J S, SinghK, Prasad MS, UmakanthB, Ram KumarG, Viraktamath BC, Madhav MS. 2013. Identification and mapping of new genetic resources for durable blast resistance in India. In: Sustainable Rice Production and Livelihood Security: Challenges and Opportunities. Cuttack: Central Rice Research Institute. [Cited within:1]
[46]
RamkumarG, SrinivasaraoK, Mohan KM, SudarshanI, Sivaranjani A KP, GopalakrishnaK, Neeraja CN, Balachand ran SM, Sundaram RM, Prasad MS, Shobha RaniN, Rama Prasad AM, Viraktamath BC, Madhav MS. 2011. Development and validation of functional marker targeting an InDel in the major rice blast disease resistance gene Pi54 (Pikh). Mol Breeding, 27(1): 129-135. [Cited within:1]
[47]
Rangel PN, Brondani R PV, Rangel P HN, BrondaniC. 2008. Agronomic and molecular characterization of introgression lines from the interspecific cross Oryza sativa (BG90-2) × Oryza glumaepatula (RS-16). Genet Mol Res, 7(1): 184-195. [Cited within:1]
[48]
Ratna MK, Srinivas PM, Laha GS, Madhan MK, Sheshu MM, Viraktamath BC. 2011. Combining blast and bacterial blight resistance in rice cultivar, improved Samba Mahsuri. Ind J Plant Prot, 39(2): 124-129. [Cited within:1]
[49]
Saghai M MA, Soliman KM, Jogensen RA, Allard RW. 1984. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location and population dynamics. Proc Natl Acad Sci USA, 81(24): 8014-8018. [Cited within:1]
[50]
Sharma TR, Madhav MS, Singh BK, ShankerP, Jana TK, DalalV, Pand itA, SinghA, GaikwadK, Upreti HC, Singh NK. 2005. High-resolution mapping, cloning and molecular characterization of the gene of rice, which confers resistance to rice blast. Mol Genet Genom, 274(6): 569-578. [Cited within:1]
[51]
SiéM, SanniK, FutakuchiK, MannehB, Mand éS, VodouhéR, DogbeS, Dramé KN, OgunbayoA, Ndjiondjop MN, TraoreK. 2012. Towards a rational use of African rice (Oryza glaberrima Steud. ) for breeding in Sub-Saharan Africa. Genes, Genom Genomics, 6(1): 1-7. [Cited within:1]
[52]
TsunematsuH, Yanoria M JT, Ebron LA, HayashiN, AndoI, KatoH, ImbeT, Khush GS. 2000. Development of monogenic lines for rice blast resistance. Breeding Sci, 50: 229-234. [Cited within:1]
[53]
Zhang FT, Xie JK. 2014. Genes and QTLs resistant to biotic and abiotic stresses from wild rice and their applications in cultivar improvements. In: Yan W G, Bao J S. Rice: Germplasm, Genetics and Improvement. Croatia, European Union: InTech. [Cited within:1]
[54]
Zhou YL, Xie XW, ZhangF, WangS, Liu XZ, Zhu LH, Xu JL, Gao YM, Li ZK. 2014. Detection of quantitative resistance loci associated with resistance to rice false smut (Ustilaginoidea virens) using introgression lines. Plant Pathol, 63(2): 365-372. [Cited within:1]
2
1992
0.0
0.0
... Though wild species have been used extensively for other important agronomic traits of rice, they have been rarely used in blast resistance breeding programmes, since only two resistance genes, Pi9 and Pi40, were reported from wild species (Amante et al, 1992 ...
... However, till today only two genes (Pi9 and Pi40) have been identified from wild species (Amante et al, 1992 ...
1
2008
0.0
0.0
1
2006
0.0
0.0
... The strategic phenotyping for blast disease (field) resistance coupled with genotyping using markers has led to the identification of resistant ILs which are valuable genetic resources not only for blast resistance but also for novel gene identification leading to cultivar development (Ashikari and Matsuoka, 2006 ...
2
2008
0.0
0.0
... In silico analysis of target regionThe introgressed regions in the four ILs were analyzed based on the existing literature for the presence of meta QTLs (Ballini et al, 2008) ...
... In addition, we observed the presence of meta QTLs in the introgressed regions of all ILs (Ballini et al, 2008), indicating the importance of these genomic regions in the point of field resistance ...
1
2003
0.0
0.0
1
1986
0.0
0.0
... oryzae have been developed, the resistance is not long lasting, because the high pathogen plasticity in the fields makes single resistance gene break down after three to five years of the cultivar release (Bonman et al, 1986 ...
1
1997
0.0
0.0
... nivara (Brar and Khush, 1997), QTLs for bacterial leaf blight and brown planthopper resistance (Zhang and Xie, 2014), and yield enhancing QTLs from various wild species (Linh et al, 2008 ...
1
2008
0.0
0.0
... DISCUSSIONWild species are rich resources for a variety of important genes which have agronomical importance (Cheema et al, 2008 ...
1
2009
0.0
0.0
... Chen et al, 2009) ...
1
2014
0.0
0.0
... To date, around 100 blast resistance genes have been identified, and many of them have been cloned and characterized, such as Pb1, Pia, Pib, Pid2, Pid3, Pik, Pik-h/Pi54, Pik-m, Pik-p, Pish, Pit, Pita, Piz-t, Pi1, Pi2/Piz-5, Pi5, Pi9, Pi21, Pi25, Pi36, Pi37, Pi35, Pi64, Pi56, Pi63 and PiCO39 (Devanna et al, 2014) ...
1
2004
0.0
0.0
1
2006
0.0
0.0
1
2008
0.0
0.0
1
2009
0.0
0.0
... Rahman et al, 2011) and also for exploring the agronomic traits (Fujita et al, 2009 ...
1
2001
0.0
0.0
... Since, field resistance allows effective control of a pathogen under natural field conditions and it is considered to be durable when exposed to new races of blast (Fukuoka and Okuno, 2001) ...
1
2014
0.0
0.0
1
2014
0.0
0.0
... PR114 and Pusa 44 are the elite rice varieties with high yield and good cooking quality (Gaikwad et al, 2014) ...
1
2006
0.0
0.0
1
2012
0.0
0.0
... Gu et al, 2012) ...
2
2006
0.0
0.0
... We also made an attempt to identify the reported blast resistance (R) genes among the resistant ILs using gene based markers which are reported earlier (Hayashi et al, 2006 ...
... Apart from stringent phenotypic screening, the gene based markers were used to identify the known blast R alleles (Hayashi et al, 2006 ...
1
2010
0.0
0.0
... Though there are many reports on the leaf blast resistance genes, less information is known towards neck blast, as only one gene Pb1 has been reported so far (Hayashi et al, 2010) ...
1
2014
0.0
0.0
1
2000
0.0
0.0
1
2014
0.0
0.0
1
2014
0.0
0.0
... Ingole Kishor et al, 2014 ...
1
2006
0.0
0.0
... Nataraj et al, 2011) including disease resistance (Jena et al, 2006 ...
2
2007
0.0
0.0
... Jeung et al, 2007) ...
... Jeung et al, 2007) ...
1
2014
0.0
0.0
... Khan et al, 2014 ...
1
2013
0.0
0.0
... 108 t in 2035 (Khush, 2013) ...
1
2009
0.0
0.0
... Lang et al, 2009) ...
1
2013
0.0
0.0
... Further, stable ILs will facilitate the use of wild species derived genes/alleles in breeding programmes (Lei et al, 2013 ...
1
2008
0.0
0.0
... nivara (Brar and Khush, 1997), QTLs for bacterial leaf blight and brown planthopper resistance (Zhang and Xie, 2014), and yield enhancing QTLs from various wild species (Linh et al, 2008 ...
3
2014
0.0
0.0
... Lmam et al, 2014) ...
... Lmam et al, 2014) ...
... Lmam et al, 2014) ...
1
2012
0.0
0.0
... Similar approach was followed earlier for identification of alleles or better performing alleles from the diverse germplasm through PCR based approach (Mahender et al, 2012 ...
2
2002
0.0
0.0
... Also, the availability of many SSR markers in rice helps the breeders for crop improvement through molecular marker assisted breeding (McCouch et al, 2002) ...
... A set of 499 SSR markers (McCouch et al, 2002) which were spread uniformly across the twelve linkage groups were used ...
1
2013
0.0
0.0
... SSR markers are most popular genetic markers with high polymorphism rate, high abundance and broad distribution throughout the genome, which are inherited in a Mendelian fashion as codominant markers (Miah et al, 2013) ...
1
2011
0.0
0.0
... Nataraj et al, 2011) including disease resistance (Jena et al, 2006 ...
1
2005
0.0
0.0
... NLR-1 maintained as DRR (Directorate of Rice Research) isolate (Padmavathi et al, 2005 ...
1
2013
0.0
0.0
... All the scores were obtained according to the standard evaluation system (Supplemental Table 3) (Panguluri et al, 2013) ...
1
2009
0.0
0.0
... Prasad et al, 2009 ...
1
2011
0.0
0.0
... For this study, seven blast R genes which are being used more often in blast resistance breeding programs in India were selected (Prasad et al, 2011) ...
1
2006
0.0
0.0
2
2011
0.0
0.0
... Rahman et al, 2011) ...
... Rahman et al, 2011) and also for exploring the agronomic traits (Fujita et al, 2009 ...
1
2013
0.0
0.0
... Wild species of Oryza can be exploited to widen the gene pool of rice for biotic and abiotic stress (Ram et al, 2013) ...
1
2013
0.0
0.0
1
2011
0.0
0.0
... Ramkumar et al, 2011) ...
1
2008
0.0
0.0
... Rangel et al, 2008 ...
1
2011
0.0
0.0
... Ratna et al, 2011) ...
1
1984
0.0
0.0
... DNA extraction and identification of blast resistance genesGenomic DNA was isolated using modified cetyltrimethyl ammonium bromide (CTAB) method (Saghai et al, 1984) and the quality of isolated DNA was checked using 0 ...
1
2005
0.0
0.0
1
2012
0.0
0.0
1
2000
0.0
0.0
... ILs have proven to be potential genetic resources for detecting rice blast resistance (Tsunematsu et al, 2000 ...
1
2014
0.0
0.0
... nivara (Brar and Khush, 1997), QTLs for bacterial leaf blight and brown planthopper resistance (Zhang and Xie, 2014), and yield enhancing QTLs from various wild species (Linh et al, 2008 ...
1
2014
0.0
0.0
... Zhou et al, 2014) ...
Development and Identification of Novel Rice Blast Resistant Sources and Their Characterization Using Molecular Markers
[S. J. S. Rama Devi1, Kuldeep Singh2, B. Umakanth1, B. Vishalakshi1, P. Renuka1, K. Vijay Sudhakar1, M. S. Prasad3, B. C. Viraktamath1, V. Ravindra Babu1, M. S. Madhav1]





{kind=link}
{kind=link}
 ]
]